Beta-1,3/1,6-D-glucan (yeast d glucan)mechanical
- Foody Bio Tech

- Sep 19, 2024
- 2 min read
As humans cannot metabolize the β-glycosidic bonds from β-glucans, it has long been suspected that the bacterial fermentation process taking place within the intestinal system is involved in the health promoting effect of β-glucans.
Meanwhile, different possible mechanisms have been identified on how oral β-glucans modulate the immune system (for review see [10,11]).
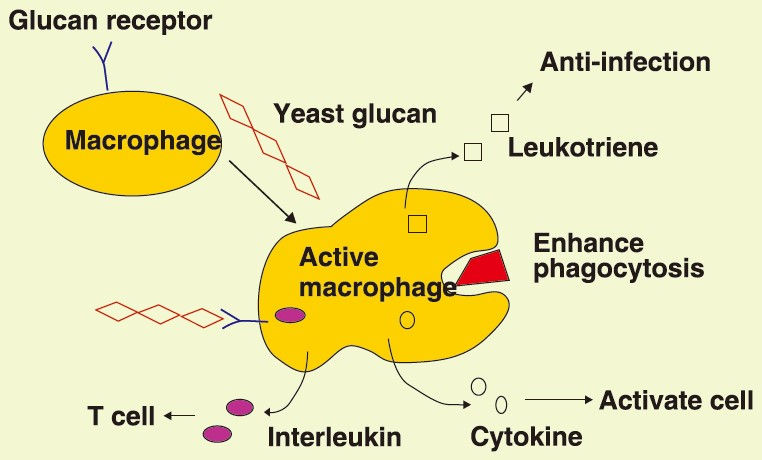
In general, humans cannot synthesize β-glucans. Therefore, the immune system recognizes these compounds as foreign. The innate immune system responds to invading pathogens through pattern recognition receptors (PRR),which are typically expressed by immune cells but also by other cells. PRRs recognize conserved microbial structures, the so-called microbe-associated molecular patterns (MAMPs) [12], formally called PAMPs [13,14]. β-glucans are considered as one of the major MAMPs for the PRR mediated sensing of fungal infection. So far, the most important PRRs for β-glucans are the dectin-1 receptor, the complement receptor 3 (CR3) and toll-like-receptors (TLR), which are found on various immune cells such as monocytes, macrophages, dendritic cells, neutrophils, eosinophils, and natural killer cells, but also on intestinal epithelial cells [10,15-17]. Binding of β-glucans to dectin-1 induced a cascade of innate and adaptive immune response such as phagocytosis, oxidative burst, and the production of cytokines and chemokines in dentritic cells and macrophages [15]. Kankkunen et al. showed that particulate yeast β-glucan triggered interleukin-1β (IL-1β) mediated cellular response in human primary macrophages via dectin-1 signaling [18]. Earlier in vitro studies showed that yeast β-glucan is a strong stimulant of macrophages [19] and induced mitogenic activity in rat thymocytes, indicating immunostimulatory effects [20].
References
10. Chan GC, Chan WK, Sze DM: The effects of beta-glucan on human immune and cancer cells. J Hematol Oncol 2009, 2:25.
11. Volman JJ, Ramakers JD, Plat J: Dietary modulation of immune function by beta-glucans. Physiol Behav 2008, 94:276–284.
12. Kumar H, Kawai T, Akira S: Pathogen recognition by the innate immune system. Int Rev Immunol 2011, 30:16–34.
13. Ausubel FM: Are innate immune signaling pathways in plants and animals conserved? Nat Immunol 2005, 6:973–979.
14. Didierlaurent A, Simonet M, Sirard JC: Innate and acquired plasticity of the intestinal immune system. Cell Mol Life Sci 2005, 62:1285–1287.
15. Brown GD, Herre J, Williams DL, Willment JA, Marshall AS, Gordon S: Dectin-1 mediates the biological effects of beta-glucans. J Exp Med 2003, 197:1119–1124.
16. Rice PJ, Adams EL, Ozment-Skelton T, Gonzalez AJ, Goldman MP, Lockhart BE, Barker LA, Breuel KF, Deponti WK, Kalbfleisch JH, Ensley HE, Brown GD, Gordon S, Williams DL: Oral delivery and gastrointestinal absorption of
soluble glucans stimulate increased resistance to infectious challenge. J Pharmacol Exp Ther 2005, 314:1079–1086.
17. Ross GD, Cain JA, Myones BL, Newman SL, Lachmann PJ: Specificity of membrane complement receptor type three (CR3) for beta-glucans. Complement 1987, 4:61–74.
18. Kankkunen P, Teirila L, Rintahaka J, Alenius H, Wolff H, Matikainen S: (1,3)-beta-glucans activate both dectin-1 and NLRP3 inflammasome in human macrophages. J Immunol 2011, 184:6335–6342.
19. Seljelid R, Bögwald J, Lundwall A: Glycan stimulation of macrophages in vitro. Exp Cell Res 1981, 131:121–129.
20. Sandula J, Machnova E: Mitogenic activity of particulate yeast b-(1,)-Dglucan and its water-soluble derivates. Int J Biol Macromol 1995,17:323–327.



Comentários